The structure of DNA
Deoxyribonucleic acid or DNA is generally found in all organisms as well as humans. DNA is an inherited material that every person has, and is set up in both the cell nucleus and the mitochondria. A higher percentage of human DNA comprise of nuclear DNA because it is found in the nucleus. (Berger, 1998). DNA comprises of numerous properties, the most significant being; that it can copy itself. In the occurrence of this process, the twofold helix strands of DNA are employed as a model for replicating the base sequences. (Watson, 2011). This is an important process in the event of cell division due to the vital requirement for each new cell to carry an accurate DNA copy found in the mature cell. Adenine (A), guanine (G), thymine (T), cytosine (C) and uracil (U) are major five chemical bases that accumulate DNA’s coded information (Watson & Crick, 2003). There are over three billion of such chemicals. More than 99% of these bases have been proved to be reliable in all humans (Buchini & Leumann, 2003). As a result of the complexity and association of the entire DNA structure, an in-depth look of each part; DNA is the genetic information of nearly all living organisms. It can be copied over age brackets of cells; it can be converted into proteins; and can be mended when required. DNA is a polymer, composed of nucleotides (Watson & Crick, 2003)
Hydrogen bonds: bases pairing
The (hydrophobic) bases are piled on the in the interior, their level surface are vertical to the axis of the double helix (Berger, 1998). The exterior (phosphate and sugar) is hydrophilic. Hydrogen limits between the bases of one strand and that of the other strand grip the two strands together (dashed lines in the drawing).
A purine on one strand links to a pyrimidine on the other strand. Consequently, the number of purines deposits matches the number of pyrimidine deposits (Watson, 2011). A binds T (with 2 hydrogen bounds), while G binds C (with 3 hydrogen bounds: more stable link: 5.5 kcal vs 3.5 kcal) (Rädler & Safinya, 1997). Thus, the substance in A in the DNA is equal to the one in T, and the substance in G equals the one in C. The complement of the 2 strands is as a result of this (A<->T and G<->C) correspondence. One serves as a template of the other, and vice versa. This feature allows exact replication (“semi-conservative replication: one strand -the template- is conserved, another is newly synthesized, same with the second strand, conserved, allowing another one to be newly synthesized”) (Buchini & Leumann, 2003).
The model of Watson and Crick above described sometimes differs from the Hydrogen bounds in base pairing from using the N7 atom of the purine instead of the N1 (Hoogsteen model).
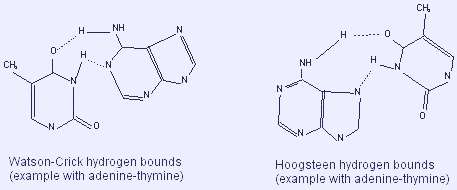
Figure 1
Major groove and minor groove
The double helix is a rather inflexible and thick molecule of a huge extent and a small diameter. It presents both major groove and a minor groove (Patil, Rhodes & Burgess, 2004). The major groove is profound and broad; the minor groove is thin and shallow. DNA-protein connections are essential procedures in the life of the cell life. Proteins connect at the ground of the DNA grooves, using a precise binding: hydrogen bounds, and distracted binding: van der Waals exchanges. Proteins recognize H-bond donors, H-bond acceptors, methyl groups (hydrophobic), which are later in the major groove (Rädler & Safinya, 1997). The major groove involves 4 likely patterns of recognition, and 2 with the minor groove. A few proteins bind DNA in its major groove, some other in the minor groove, and some need to bind to both.
The minor groove of double helical B-DNA is a sector of great attention for rising new drugs because of its non-covalence high succession specific connections for a huge number of tiny molecules (Berger, 1998). Minor groove binding lies among the broadly studied class of agents exemplified by, an advanced succession specificity and possessing diverse biological actions. A number of them display antiviral, antibacterial, and antiprotozoal properties. However, others have shown antitumor activity.
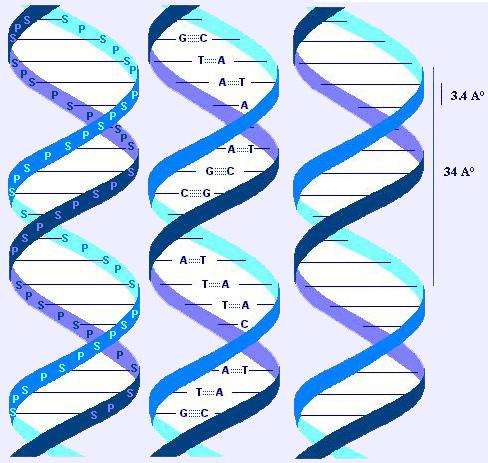
Figure 2
H-DNA or triplex DNA
Inverted repeats (palindromes) of polypurine/polypyrimidine DNA stretches can structure triplex structures (triple helix) (Watson & Crick, 2003). A triple-stranded together with a single stranded DNA are formed. H-DNA may have a function in practical regulation of gene appearance as well as on RNAs (e.g. repression of transcription).
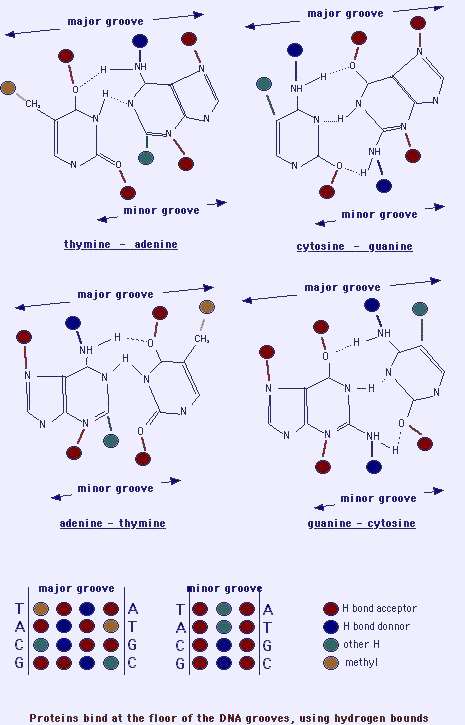
Figure 3
Triplex Forming Nucleotides
Triplex forming oligonucleotides (TFOs) have attained significant focus as a possible therapeutic agent to aim gene expression (Patil, Rhodes & Burgess, 2004). They are a group of DNA oligonucleotides which are capable of fusing with other main groove of the duplex DNA creating triple helix (Buchini & Leumann, 2003). The creation of a triplex beside the main groove leads to competition with the fusing of transcription factors and the proteins essential for transcription. TFOs provide specificity sequence and hence can be used to aim and inhibit appearance of specific genes which are associated with a particular disease state. In addition TFOs can also be used as diagnostic agents for identification of a foreign DNA (viral or bacterial) or any diseases associated to mutations.
Binding of Triplex Forming Oligonucleotides
TFOs, in the span of approximately 20 bases, can bind in the major groove via Hoogsteen hydrogen bonds to the purine (A and G) bases on the double stranded DNA, already in the structure of the Watson-Crick helix. The binding can take place at the extent of pyrimidines of one DNA strand and the opposite purines on the other. TFOs bind to the strand with purines.
Respective of their base composure, TFOs can bind to the double helix in either parallel or antiparallel direction to the purine-bearing strand (Buchini & Leumann, 2003). TFOs made of pyrimidines (C and T) bind to the purine-rich strand of the objective double helix via Hoogsteen hydrogen bonds in a parallel manner. TFOs comprised of purines (A and G), or mixed purine and pyrimidine (G and T) bind to the same purine-rich strand through reverse Hoogsteen bonds in an anti-parallel style (Rädler & Safinya, 1997).
Oligonucleotides with modified structural features
Oligonucleotides hauling other compound moieties besides the normal bases are called “modified oligonucleotides”. The modifications can be situated at the 3’- or 5’-end of the oligonucleotide, or within the sugar-phosphate stamina or at the nucleobases. Mainly all PCR applications are still potential if the modification is close to the 5’- end of the oligonucleotide. Contrary, modifications at the 3’- end typically wedge this end for extra enzymatic response. In order to achieve an absolute blocking, an inverted end or C3-Spacer modification is recommended (Patil, Rhodes & Burgess, 2004). Chemically synthesized oligonucleotides bear free hydroxy (OH) collection at their relevant 3’- and 5’- ends (Berger, 1998). Besides, some biological tests need the presence of the natural structure of an oligonucleotide. This modification requires being ordered explicitly (Rädler & Safinya, 1997).
Structure and Limitations
DNA triple helices figure in a sequence-specific way on polypurine:polypyrimidine tracts (1–3), which are extensive in mammalian genomes (4–6). The third thread recline in the major groove of an integral duplex and is calmed by two Hoogsteen hydrogen bonds between third strand bases and the purines in the duplex (3, 7) (Buchini & Leumann, 2003). The third strand may comprise of pyrimidines, or purines, respective of the character of the target succession. In the pyrimidine (or Y.R:Y) motif, a homopyrimidine oligonucleotide binds in a trend parallel to the purine strand in the duplex, with canonical base triplets of T.A:T and C.G:C. In the alternate purine motif (R.R:Y), a homopurine strand binds antiparallel to the purine strand, with base triplets of A.A:T and G.G:C (8, 9) (Buchini & Leumann, 2003). The development of TFOs could involve sequence-specific gene targeting reagents in live cells (12–17) (Berger, 1998).
Despite this, several obstacles still need to be overcome. Triplex chemistry and biochemistry inflict essential limitations to TFO action in the nuclear setting, and target choices are inadequate to polypurine:polypyrimidine sequences (Watson, 2011). Additionally, it is evident that nucleosomes can inhibit triplex formation (18–20). As a result, ease of access to genomic targets is an important issue.
Potential applications of TFOs-gene targeting
Potential applications of TFOs embrace gene targeting treatment particularly for cancer and the study of gene expressions. TFOs can hush a gene record by aiming the dictation initiation sites, (i.e., the promoter region), or by targeting recording of elongation by striking at the triplex binding sites. Specifity of sequence is the answer to efficient genetic targeting. With the use of specificity, genes that are targeted can be changed in many ways. Gene therapy agents change into loose cannons inside the cells without it. Triplex forming oligonucleotides (TFOs) fuse into main groove of the duplex DNA with high affinity and specificity (Watson, 2011). Due to these properties, TFOs have been thought as host devices for the genetic manipulation. Recent researches have shown that TFOs have the ability to mediate targeted gene success in mice, establishing the basis for the possible application of those molecules in a human being’s gene therapy.
Molecules that fuse with the DNA double helix might interfere with gene appearance and, to add to the potential therapeutic applications, it can be useful for the research of DNA processing, package of chromatin, or related biological processes. The Triplex-forming oligonucleotides (TFOs) fuse with specific sequences in DNA double helix through hydrogen bonding interactions. The TFOs have been revealed to down-regulate the expression of the gene, to induce aimed genomic DNA modifications, so as to fasten DNA combination, and also to regulate chromatin organization. In addition to this, they can be used as transporting agents to place DNA-modifying agents into selected sequences (Patil, Rhodes & Burgess, 2004). Something important regarding TFO technologies are the creation of fresh oligonucleotide analogues which have improved fusing affinity, sufficient stability and better target selectivity, in intracellular environment.
References
Watson, J. D, & Crick, F. H, 2003, The structure of DNA. In Cold Spring Harbor Symposia on Quantitative Biology, Vol. 18, pp. 123-131, Cold Spring Harbor Laboratory Press.
Berger, J. M. (1998). Structure of DNA topoisomerases. Biochimica et Biophysica Acta (BBA)-Gene Structure and Expression, 1400(1), 3-18.
Rädler, J. & Safinya, C. R., 1997, Structure of DNA-cationic liposome complexes: DNA Intercalation in Multilamellar Membranes in Distinct Interhelical Packing Regimes, Science, 275(5301), 810-814.
Watson, J. D, 2011, The double helix: A personal account of the discovery of the structure of DNA, Simon and Schuster.
Buchini, S., & Leumann, C. J, 2003, Recent improvements in antigene technology, Current opinion in chemical biology, 7(6), 717-726.
Patil, S. D., Rhodes, D. G., & Burgess, D. J, 2004, Anionic liposomal delivery system for DNA transfection, The AAPS journal, 6(4), 13-22.
You have to be 100% sure of the quality of your product to give a money-back guarantee. This describes us perfectly. Make sure that this guarantee is totally transparent.
Read moreEach paper is composed from scratch, according to your instructions. It is then checked by our plagiarism-detection software. There is no gap where plagiarism could squeeze in.
Read moreThanks to our free revisions, there is no way for you to be unsatisfied. We will work on your paper until you are completely happy with the result.
Read moreYour email is safe, as we store it according to international data protection rules. Your bank details are secure, as we use only reliable payment systems.
Read moreBy sending us your money, you buy the service we provide. Check out our terms and conditions if you prefer business talks to be laid out in official language.
Read more